About this pathway
Background
Carbamazepine (CBZ), a dibenzazepine, is a tricyclic compound used in the treatment of epilepsy, trigeminal neuralgia and psychiatric disorders [Article:18463198]. Serious adverse events have been reported for CBZ including Stevens-Johnson Syndrome (SJS), Toxic Epidermal Necrolysis (TEN) and drug reaction with eosinophilia and systemic symptoms (DRESS) [Articles:20235791, 18717872]. Other types of hypersensitivity reactions are also associated with CBZ including mild skin rashes, fever, eosinophilia, and cross-reactions to other anticonvulsants. Up to 80% of patients who have an idiopathic drug reaction to CBZ drugs will also have an adverse reaction to other anticonvulsants, further restricting treatment options [Article:3198757]. In addition to adverse events, lack of efficacy can also be a problem, with as many as 30% of patients with epilepsy experiencing drug-resistance [Articles:18088268, 19415824]. The mechanisms by which these events occur are not entirely clear although several candidate pharmacogenes have been associated with CBZ treatment responses. Current methods to individualize treatment involve therapeutic drug monitoring, the measurement of drug metabolites in patient samples post-treatment and subsequent dose adjustment. While this provides an accurate view of the drug response phenotype it still risks adverse events and cross-sensitivity. The ability to identify the patients that will benefit from CBZ, not suffer adverse events and define dose prior to treatment would be a highly valuable clinical tool. Here we present the current knowledge of CBZ pharmacogenomics (PGx) as a gene centered view of the pharmacokinetics of CBZ and collate the gene variants associated with CBZ responses it.
Pharmacokinetics
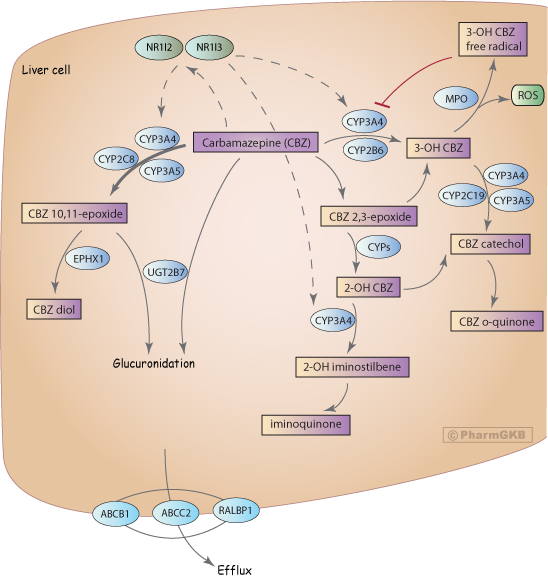
CBZ is almost completely metabolized in the liver with only around 5% of the drug excreted unchanged [Article:15915352]. The major route of metabolism is conversion to CBZ 10.11-epoxide (CBZ-E)[Article:18463198]. This reaction is primarily catalyzed by CYP3A4 although CYP2C8 also plays a role, and involvement of CYP3A5 has also been suggested [Articles:18463198, 8010982]. Minor metabolic pathways include ring-hydroxylation to form 2-hydroxy-CBZ (2-OH-CBZ) and 3-hydroxy CBZ (3-OH CBZ). The formation of each presumably proceeds via an epoxide intermediate (referred to as an arene oxide intermediate), with CYP2B6 and CYP3A4, being the major catalysts of 3-OH-CBZ formation [Article:18463198], and multiple CYPs involved in 2-OH-CBZ formation [Article:12386121]. Secondary metabolism of 2-OH-CBZ and 3-OH-CBZ by CYP3A4 represent two distinct potential bioactivation pathways. CYP3A4-dependent secondary oxidation of 2-OH-carbamazepine leads to the formation of thiol-reactive metabolites via an iminoquinone intermediate [Article:16135660], whereas CYP3A4-dependent secondary oxidation of 3-OH-CBZ results in the formation of reactive metabolites capable of inactivating CYP3A4 [Article:18463198] and forming covalent adducts [Article:18096676]. 3-OH CBZ, and to a lesser extent 2-OH CBZ and CBZ, can be metabolized to form radicals by myeloperoxidase (MPO)[Article:18463199]. This releases reactive oxygen species and may lead to the formation of protein adducts. Covalent binding and protein adduct formation has also been observed for another anti-epileptic drug, phenytoin, and is generally considered to be a necessary step in the pathogenesis of idiosyncratic reactions to this class of compounds [Article:18463199].
CBZ stimulates the transcriptional upregulation of genes involved in its own metabolism, with autoinduction of CYP3A4 and CYP2B6, via NR1I2 (PXR) and NR1I3 (CAR) [Articles:3319544, 17112801, 19702527]. Drug-drug interactions via CYP3A4 [Article:10668858] and CYP2B6 [Article:19223624] are well documented and can complicate the use of CBZ in polytherapy.
Some studies have suggested that glucuronidation is likely to be only a minor role in metabolism of CBZ and CBZ-E [Article:15915352]. But other papers dispute that documenting involvement of UGT2B7 [Articles:15292462, 17495417].
Transport
Variable transport of CBZ, particularly across the blood brain barrier, may be responsible for variable CBZ response. Increased export from the brain has been discussed as a method of drug resistance with P-glycoprotein (PgP, coded for by ABCB1) as the main focus [Article:18088268]. Although studies in rats suggested PgP transport of CBZ [Article:11733711], in vitro assays and work in mice did not show evidence of CBZ transport by PgP [Articles:11318771, 18824002]. Recent work has confirmed CBZ is not transported by PgP although it did transport CBZ 10, 11-epoxide [Article:21692796]. RALBP1, also known as RLIP76, has been shown to transport CBZ and be involved in drug resistance [Article:16188027]. Additional PGx evidence implicates ABCC2 as a potential pharmacogene for CBZ [Article:20216337] however cellular studies showed ABCC1, ABCC2 and ABCC5 did not transport CBZ in vitro [Article:20080116]. See below for discussion of specific genomic variants in transporters and CBZ PGx.
Pharmacogenomics
Major Histocompatibility Locus Variants
The most well studied PGx variants with respect to CBZ are variants within major histocompatibility (MHC) locus in the human leukocyte antigen gene HLA-B [Article:18370849]. HLA-B codes for a protein that presents peptides to the immune system, identifying foreign or infected cells [Article:518865]. There are over 1500 alleles of HLA-B according to the IMGT/HLA Database linkout[Article:21071412]. Historically these were identified by serotype phenotyping and although sequencing is now common, the definitive genomic variants associated with these are not well described. The extreme diversity of this locus in different ethnic groups means that different tag SNPs are associated with the different serotypes in different populations [Article:16998491]. The allele most associated with risk for the severe ADRs, SJS and TEN, in response to CBZ is HLA-B*1502 [Article:15057820]. One mechanism that has been suggested for how CBZ hypersensitivity is triggered involves the proteasomal degradation and MHC-dependent presentation of CBZ metabolites [Article:18991696]. The generation of free radicals is considered another possible mechanism. By forming adducts with CYP3A4 enzyme the radicals may also contribute to the cross-reactive hypersensitivity sometimes seen with other antiepileptic drugs [Article:18096676]. An in vitro study showed covalent binding of CBZ-modified peptides to the HLA-B*1502 protein that may lead to T-cell activation and SJS with this allele specifically [Article:17697703].
HLA*1502 allele is strongly associated with CBZ-induced SJS/TEN in Taiwanese, Chinese, Indians and Chinese-Americans but not in Caucasians or Japanese individuals [Articles:15057820, 16538176, 20833111, 19915237, 16415921, 16981842, 18637831, 19018717]. This has led the clinical labeling from the FDA to recommend testing only in individuals with ancestry in genetically at risk populations FDA linkout. The tag SNPs for HLA-B*1502 in Han Chinese HapMap samples are SNPs rs3909184 and rs2844682 [Article:16998491].
HLA-A*3101 has been associated with CBZ-induced ADRs in Asians [Article:16538176]. Recently two independent GWAS studies showed association of HLA-A*3101 with CBZ-induced ADRs in Caucasians [Article:21428769] and Asians [Article:21149285]. The tag SNP in linkage with HLA-A*3101 in the Asian population was rs1633021 [Article:21149285] and in the Caucasian population was rs1061235 [Article:21428769].
Due to high degree of linkage across the MHC region tagging SNPs may tag for a functional variant in another gene. The variants rs3909184 (within FLOT1 gene), rs2844682 (MUC21), rs1059510 (HLA-E), rs1264511 (intergenic), rs3130690 (intergenic), rs2848716 (intergenic), rs750332 (BAT2), rs2227956 (HSPA1A, HSPA1L, LSM2), rs1043620 (HSPA1A, HSPA1L, LSM2), rs506770 (HSPA1A, HSPA1L), rs2395402 (LEMD2) rs986475 (LST1, LTB, NCR3), rs2894342 (MLN) and rs1800629 (TNF:(-308)G>A) have been associated with CBZ-induced ADRs [Articles:16538176, 16538175, 11294926]. Also, the HLA-B*0702 allele was shown to protect against severe CBZ hypersensitivity in a small study of Caucasians [Article:16981842]. Since not all individuals with the HLA*1502 allele experience ADRs it is still unclear which particular SNPs are causative and which are just tagging SNPs. The definition of which SNPs are causative will aid in better identifying those patients at risk for ADR particularly in those without Asian ancestry.
Metabolizing Enzyme Variants
Variants in CBZ metabolizing enzymes have been shown to affect CBZ pharmacokinetics although studies have been few and without replication. The reduced function protein CYP3A4*16 (rs12721627) shows decreased clearance in in vitro systems [Articles:19255940, 20847137] therefore potentially requiring altered dosing in individuals with this variant (found at a frequency of 1-5% in populations from Japan, Korea and Mexico). Clearance of CBZ may be altered in vivo by CYP3A5 variants [Articles:19744012, 16678552]. A small study of Korean individuals with epilepsy found that CYP3A5 non-expressers (CYP3A5*3, rs776746) had higher clearance of CBZ as well as higher plasma levels than CYP3A5 expressers, a finding that seems incongruent but could be explained by autoinduction of CYP3A genes [Article:19744012]. However a larger study of Japanese epilepsy patients did not find a difference although this study included patients on co-medications that may have further induced CYP3A4 [Article:16678552]. Variants in EPHX1 have also been associated with altered CBZ metabolism [Article:15692831]. A haplotype of rs1051740 (EPHX1: Y113H) and rs2234922 (EPHX1: H139R) showed increased plasma CBZ-diol/CBZ-epoxide ratios in vivo in Japanese epilepsy patients [Article:15692831]. Studies of polymorphisms in metabolizing drugs and effect on CBZ-induced ADRs have been mostly negative [] with one study associating a SNP in the 3'UTR of CYP2B6 (rs1042389) with maculopapular eruption and hypersensitivity syndrome but this was not significant after Bonferroni correction [Article:16538176].
Transporter Variants
Work on the PGx of CBZ transport and resistance are similarly conflicting and in need of replication in larger cohorts. The well-known ABCB1 variant 3435C>T, rs1045642 CC genotype was associated with drug-resistant epilepsy in a cohort of 315 British patients although the drugs used in this study were not specified [Article:12686700]. Several studies since then have found no association of this variant (see meta-analysis by Bournissen et al) and these too did not separate patients by treatment [Article:19178561]. A study of 464 Chinese epilepsy patients associated ABCB1 variants rs3789243 and rs2032582 with CBZ resistance [Article:19450124], but a study of 228 North Indian epileptics did not replicate this association [Article:20417680]. While it could be that different haplotype structures or racial background may have influenced these results, a sub-analysis of the Bournissen meta-analysis which looked at European cohorts and Asian cohorts separately also found no evidence of association of ABCB1:3435C>T with drug resistance [Article:19178561].
Initial studies of RALBP1 expression pointed towards a role in drug resistant epilepsy [Article:16188027] however two studies of RALBP1 variants in British cohorts failed to find association for all treatments [Articles:17437410, 18086001]. There was a weak association in the small subset of patients (n = 81) on CBZ only for rs329017 where the p values were not significant but the authors felt warranted further study [Article:18086001]. One association that has been replicated is with the ABCC2 SNP c.1249G>A (p.V417I, rs2273697). This variant was associated with neurological ADRs in 146 Korean individuals with epilepsy receiving CBZ and validated in an independent cohort of the same ethnicity [Article:20216337] An additional SNP in ABCC2 (-24C > T, rs717620) has been associated with lack of response to CBZ in young Caucasian epilepsy patients [Article:19415824].
Pharmacodynamic Variants
Although not depicted in the figure, the targets of CBZ the brain sodium channels SCN1A, SCN1B, SCN2A and SCN3A have pharmacogenomic consequences. The variant SCN1A IVS5N+5 G>A (rs3812718, also reported as SCN1A IVS4-91G>A) has been associated with high dose requirements in epilepsy patients [Articles:15805193, 17436242]. Variants in SCN2A and SCN3A may contribute to CBZ resistance [Articles:18784617, 18242854] in individuals with epilepsy. In vitro evidence from mice also suggests SCN1B as a potential pharmacogene for CBZ that may warrant further study [Article:20573896].
Conclusions
The HLA alleles (HLA-B*1502 and HLA-A*3101) are the most important pharmacogenomic variants for carbamazepine to date. While it is encouraging that labeling changes have been made for CBZ that have been shown to prevent severe side effects [Article:21428768], we still need to understand the mechanism by which these events occur and how ethnicity influences this so as to test more reliably in individuals of unknown or mixed ancestry.
Although preliminary data has been collected to show influence of genomic variation on CBZ metabolism studies have been small and not validated. Work on defining the PGx of drug resistance has been complicated by common cotreatment with several antiepileptic drugs. There is the need for larger studies that have sufficient numbers in each of the documented treatment groups with well-defined phenotypes. More work is needed to translate observed differences in metabolism and pharmacokinetics into using genomic variation for predictive dosing.
Edit history (3)
- 2011-02-14 Create
- 2011-11-16 Update
- 2024-07-08 Update fixed typos