About this pathway
Introduction
Pegylated interferon-alpha (Peginterferon-alfa or PEG-IFN-α) is an antiviral and immunoregulatory drug that has served as the foundation for treatment of hepatitis C virus (HCV) infection for more than two decades. Currently, there are two PEG-IFNs on the market in combination with the broad-spectrum antiviral, ribavirin (RBV), for treatment of chronic HCV: PEG-IFN-α 2a and PEG-IFN-α 2b. The two drugs are differentiated by the size and nature of their covalently attached polyethylene glycol (PEG) moiety. While PEG-IFN-α 2a and 2b exhibit resulting differences in pharmacokinetics and dosing regimes, the consensus is that the two drugs are clinically equivalent [Article:20108989].
HCV infection affects 170-200 million people worldwide and is the leading cause of cirrhosis and hepatocellular carcinoma (World Health Organization. Hepatitis C. WHO fact sheet 164. Geneva, Switzerland: World Health Organization; 2000. Accessed April 28, 2014. http://www.who.int/mediacentre/factsheets/fs164/en/). Although the prevalence of HCV appears to be decreasing in most developed nations, mortality due to liver disease secondary to HCV infection is expected to continue to rise over the next 20 years [Article:23280550]. While a minority of patients with acute HCV infections are able to clear the virus spontaneously, 55-85% of patients develop chronic HCV infection, defined as detectable HCV RNA for longer than 6 months [Article:12407573]. Chronic infection, once established, rarely resolves spontaneously [Article:17151366]. The primary goal for treatment is achievement of sustained virologic response (SVR), characterized by undetectable serum viral RNA 24 weeks after the end of therapy, which is considered clinically as cure of infection and is associated with lower morbidity and mortality [Article:23268517]. HCV is divided into seven recognized genotypes (1-7), which differ at 30-35% of nucleotide sites [Article:8175159]. HCV genotypes are differentially distributed globally, with genotype 1 being most prevalent in North America, South America and Europe [Article:25069599]. Further, HCV genotype is highly associated with response to PEG-IFN-α/RBV combination therapy and rates of treatment-induced SVR are lowest for patients infected with HCV genotypes 1 and 4 [Article:19330875].
Prior to 2011, the standard-of-care for patients with chronic HCV has been a combination of PEG-IFN-α/RBV therapy (either PEG-IFN-α 2a or 2b) administered for either 48 weeks for HCV genotypes 1, 4, 5, and 6, or 24 weeks for genotypes 2 and 3. However, these combination therapies yield SVR rates of only 40-50% in HCV genotype 1 patients [Article:21696309]. In contrast, SVR rates for genotypes 2 and 3 are approximately 70-80%, although these apply to selected populations without the co-morbidities that often accompany HCV [Articles:21696309, 11583749, 12324553, 14996676, 12407598]. Further, combination therapy is costly, associated with several moderate to severe side effects (influenza-like symptoms, depression, thrombocytopenia and hemolytic anemia) [Article:16107837], and contraindicated in many patients (hepatic decompensation, portal hypertension, hypersplenism, severe psychiatric depression, major systems impairment, and pregnancy) (World Hepatitis C: Global Alert and Response. Geneva, Switzerland: World Health Organization; 2000. Accessed February 8, 2015. http://www.who.int/csr/disease/hepatitis/whocdscsrlyo2003/en/index5.html). Positive predictive factors for achieving SVR include young age, female sex, and low pretreatment HCV-RNA levels [Article:25114601]. Conversely, SVR rates are less likely in HCV patients with high baseline HCV-RNA levels (>800,000 IU/mL), steatosis, insulin resistance, co-infection with HIV, and more advanced liver fibrosis [Articles:25114601, 17151366].
The strongest pre-treatment predictor of SVR rate is variations in IFNL3, (formerly known as IL28B) located on chromosome 19. The gene encodes IFN-lambda 3 (IFN-λ3), a member of the type 3 IFN-λ family that exhibits antiviral, antiproliferative and immunomodulatory activities [Article:20712453]. The strongest and most commonly tested polymorphisms are rs12979860 and rs8099917, which are located 3 kb and 8 kb upstream of IFNL3, respectively, and exhibit strong linkage disequilibrium [Article:22118055]. As described in detail below, patients with the favorable rs12979860 CC or rs8099917 TT genotype have greater than a 2-fold increased likelihood of achieving treatment-induced SVR. The Clinical Pharmacogenetics Implementation Consortium (CPIC), which advises clinicians on how to utilize genetic information in treatment decisions, has produced detailed guidelines based on IFNL3 genotyping for PEG-IFN-α/RBV-based treatment regimes [Article:24096968]. IFNL3 genotype can inform decisions on whether to initiate PEG-IFN-α/RBV therapy given the risk of adverse events and treatment failure. Once the decision to initiate therapy is made, IFNL3 genotype can be used to inform decisions on both treatment composition (e.g. whether to prescribe PEG-IFN-α/RBV dual therapy or triple therapies that includes DAAs) and duration (e.g. whether to shorten treatment duration in certain patients). The current consensus is that rs1297860 (merged into rs154739) genotype is the SNP most likely to predict treatment response if a single SNP has to be selected for diagnostic purposes, although tests for both rs12979860 and rs8099917 genotyping are commercially available [Articles:21254181, 24114761].
Recently, the advent of direct-acting antiviral agents (DAAs), which act to directly target specific stages of the HCV life cycle, has allowed for even higher SVR rates in HCV genotype 1 patients. DAAs act to directly target specific stages of the HCV life cycle. Although PEG-IFN and RBV remained essential components of therapy with the first waves of these medications, they are no longer part of the recommended regimens. The first regimens were called triple therapies and included one DAA in combination with PEG-IFN and RBV. These regimens significantly improved SVR rates and allowed for shorter treatment regimes in many patients [Article:21898493]. In 2011, the American Association for the Study of Liver Diseases (AASLD) updated its practice guidelines for chronic HCV genotype 1 patients to triple therapies that include either of the 'first generation' protease inhibitor DAAs boceprevir (BOC) or telaprevir (TVR) [Article:21898493]. These triple therapies have demonstrated SVR rates of 65-75% [Articles:21449784, 21696307, 21696308]. The 'second generation' protease inhibitor simeprevir was approved by the FDA in 2013 and raised SVR rates to approximately 80% [Article:25192745]. The recently FDA-approved sofosbuvir has shown great potential to improve SVR rates even further, either as triple therapies in combination with PEG-IFN-α/RBV [Articles:24266912, 23944316, 23499440] or as part of IFN-free therapies [Articles:24262278, 24209977]. Lastly, trials of the recently FDA-approved IFN-free regime consisting of paritaprevir (ABT-450) boosted by ritonavir, ombitasvir, dasabuvir and ribavirin, have demonstrated SVR rates greater than 95% in both treatment-naïve patients and previous non-responders with HCV genotype 1 and no cirrhosis [24720703, 24720679]. As a result, the AASLD recently revised its practice guidelines for HCV genotype 1 patients to include only IFN-free regimes, which boast improved tolerability as well as efficacy (American Association for the Study of Liver Diseases. Recommendations for testing, managing, and treating hepatitis C; 2014. Accessed April 28, 2014. http://www.hcvguidelines.org).
Pharmacokinetics
Attachment of a polyethylene glycol (PEG) moiety (pegylation) was a major advancement to improve pharmacokinetics of IFNs. Compared to thrice weekly administered unmodified IFN-α, PEG-IFNs allow for a once-weekly dosing administration and avoid large fluctuating serum concentrations. PEG-IFN-α 2a has a branched 40 kDa PEG chain covalently attached to lysine residues and circulates as an intact molecule, while PEG-IFN-α 2b has a linear 12 kDa PEG chain covalently attached via an unstable urethane bond that is hydrolyzed after injection. The size and nature of the PEG moiety attached causes differences in the pharmacokinetics and dosing regimens of the drugs [Article:22233411].
The absorption half-life of unmodified IFN-α is 2.3 hours, as compared to that of PEG-IFN-α 2a and 2b, which are approximately 50 hours and 4.6 hours, respectively [Article:12934165]. In addition to its longer half-life, PEG-IFN- α 2a is more highly localized than PEG-IFN- α 2b, having a smaller volume of distribution (0.99 L/kg) with highest concentrations occurring in the liver [Article:12934165]. Patients administered a single dose of PEG-IFN- α 2a reached a mean maximum serum concentration of 14.2 µg/L in a mean time of 78 hours after administration [Article:20951922]. Following multiple doses (180 µg weekly), mean maximum serum concentration was 25.6 µg/L, which was reached in a mean time of 45 hours. In contrast, maximum serum concentrations for PEG-IFN-α 2b are achieved between 15 and 44 hours after administration and are sustained for 48 to 72 hours. PEG-IFN- α 2a has a smaller peak-to-trough ratio of 1.5 to 2, as compared to >10 after multiple doses. Thus, PEG-IFN- α 2a exhibits less fluctuation in serum concentration during the one-week dosing interval [Article:12934165].
The longer half-life and limited distribution of PEG-IFN-α 2a allows for a fixed, weekly dose. Conversely, weekly dosage of PEG-IFN-α 2b must be adjusted according to body weight [Article:20108989]. Due to the relatively short serum half-life of PEG-IFN α 2b, a significant number of patients may have undetectable levels of the drug by the end of the weekly dose interval, which may cause viral rebound [Articles:15456079, 17875007]. For this reason, it has been suggested that a shorter dosage interval, such as a twice-weekly regimen, may be necessary for PEG-IFN-α 2b, although this has not yet been formally tested [Article:20951922].
PEG-IFN-α 2a is cleared by both the liver and kidney; approximately 30% of PEG-IFN- α 2b is cleared by the kidney, with the rest cleared by the liver or degraded after interacting with cellular IFN receptors [Article:12934165]. Compared to standard IFN-α, PEG-IFN-α 2a and 2b have significantly reduced renal clearance [Article:12052715].
RBV co-administration does not affect the pharmacokinetics of PEG-IFNs nor do PEG-IFNs affect that of RBV [Article:10960463]. For patients administered PEG-IFN- α 2b/RBV combination therapy, mean peak plasma RBV concentrations at week 1 were 741 ng/mL, 799 ng/mL, and 1101 ng/mL for daily RBV doses of 600, 800 and 1000-1200 mg, respectively. At week 4, these dosing regimes produced mean peak plasma RBV concentrations of 1770 ng/mL, 2297 ng/mL and 2750 ng/mL, respectively, accounting for a 5-fold RBV accumulation. Regardless of dosing group, apparent clearance of RBV was consistently 23-26 L/h [Article:12934165].
Pharmacodynamics
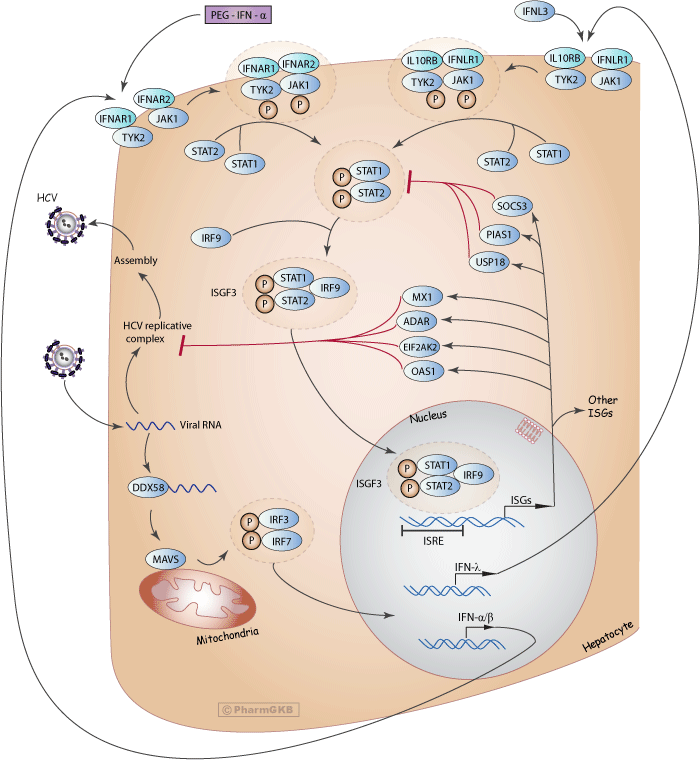
IFN-α acts both directly and indirectly as an antiviral, although the exact mechanisms by which PEG-IFN-α inhibits HCV replication are still not well understood [Article:20108989]. IFN-α exerts its effects by inducing a non-virus-specific immune response in cells through up-regulation of many IFN-stimulated genes (ISGs) that encode antiviral effector proteins. As shown in the above diagram (Figure 1), when HCV RNA is detected by the host innate immune system, it is recognized by several pathogen recognition receptors (PRRs) such as retinoic acid inducible gene-1 (RIG-1), encoded by the DDX58 gene. RIG-1 activates the adapter mitochondrial antiviral signaling protein (MAVS), which induces phosphorylation of IFN regulatory factor 3 (IRF3) and IRF7. The two proteins heterodimerize, then translocate to the nucleus and induce expression and secretion of various IFNs, including IFN-α, IFN-β and IFN-λ.
Circulating IFN-α binds to two unique cell surface receptor subunits, IFN-α R1 (IFNAR1) and IFN-α R2 (IFNAR2). Once bound to IFNAR1 and IFNAR2, a heterodimer forms and activates Janus-activated and tyrosine kinases (JAK) which in turn phosphorylate the cytoplasmic signal transducers and activators of transcription (STAT) proteins, among them STAT1 and STAT2. Phosphorylated STAT1 and STAT2 dimerize and bind IFN regulatory factor 9 (IRF9), forming the IFN-stimulated gene factor 3 (ISGF3) complex. ISGF3 then translocates to the nucleus, where it binds to IFN-stimulated response elements (ISREs) on DNA, inducing transcription of ISG mRNAs encoding a number of effector proteins.
Microarray analyses have shown that IFN-α induces hundreds of ISGs, many of which are related to antiviral activity but others involved in lipid metabolism, apoptosis, protein degradation and inflammatory responses [Article:11404376]. Among the major ISGs believed to play important roles in inhibiting HCV replication are 2',5' oligoadenylate synthetase 1 (OAS1), eukaryotic translation initiation factor 2-alfa kinase 2 (EIF2AK2, formerly known as PKR), adenosine deaminase RNA-specific (ADAR) and MX dynamin-like GTPase 1 (MX1). Conversely, antiviral signaling is controlled by the anti-inflammatory ISGs, such as ubiquitin specific peptidase 18 (USP18), the suppressor of cytokine signaling 3 (SOCS3) and protein inhibitor of activated STAT 1 (PIAS1), which act as part of a negative feedback loop that restricts both the extent and duration of the IFN response by direct or indirect inhibition of STAT signaling [Articles:22956738, 17525754].
In addition to its antiviral activity, IFN-α exerts immunologic effects through interaction with the adaptive and innate immune responses to promote memory T-cell proliferation, prevent T-cell apoptosis, stimulate natural-killer cell activation and dendritic-cell maturation [Article:9041265]. Lastly, IFN-α has been shown to increase production of major histocompatibility complex (MHC) class-I and class-II molecules [Article:10790425]. However, these immunological properties are likely not directly involved in HCV clearance with IFN-based therapies, which are thought to exert their effects primarily due to their antiviral activity [Article:23886001].
Virological decline, often referred to as the "viral kinetics of response," is an important on-treatment predictor of achieving SVR during PEG-IFN-α/RBV therapy. Rapid virological clearance (RVR) and early virological response (EVR), defined as undetectable HCV RNA after 4 weeks and a 2-log decline in HCV RNA after 12 weeks, respectively, have both been shown to be associated with SVR [Articles:21371579, 21898493]. Indeed, SVR rates for patients who achieve an RVR are greater than 70% and treatment regime can be shortened in these cases [Articles:18069698, 19780879].
The pharmacodynamics of PEG-IFN-α 2a versus 2b appear to be similar. Pharmacodynamic comparisons have shown the two PEG-IFN-α forms exhibit similar biological activity, as evidenced by induction patterns of 2',5' OAS, neopterin and the MHC class I molecule β 2-microglobulin [Articles:17635371, 20108989]. However, one study found that PEG-IFN-α 2b exhibited greater activation of ISGs in a group of 31 patients with HCV genotype 1 infection [Article:16780997].
Similar to IFN-α, IFN-λ activates the Janus kinase-signal transducer and activator of transcription (JAK-STAT) pathway, which up-regulates a number of ISGs via binding to the IFN-stimulated response element (ISRE) and establishes an antiviral state in cells. IFN-λ has been shown in experimental models to inhibit HCV replication, and when co-administered with IFN-α, this antiviral effect was additive [Articles:15731279, 17087946]. Although IFN-α and IFN-λ increase the expression of a similar set of ISGs via the same downstream pathways to exert their effects, they signal through unique receptors: IFN-λ binds to the IL10R-IL28R receptor complex, which is highly expressed only in hepatocytes [Article:12483210]. Further, the kinetics of response differs between the two IFNs: IFN-λ exhibits more rapid phosphorylation of STAT yet more delayed and prolonged expression of various ISGs compared to IFN-α [Article:17087946]. Due to the more limited distribution of the IFN-λ receptor, among other factors, it has been suggested that a pegylated form of IFN-λ may be better tolerated than PEG-IFN-α [Articles:21147189, 20564352].
The mechanism of action of RBV against HCV remains unclear. It has been suggested that RBV acts directly to inhibit HCV replication or inhibits the host enzyme inosine monophosphate dehydrogenase (IMPDH) [Article:11981750]. Conversely, RBV may act indirectly against HCV viral replication by causing rapid and lethal mutations of virions or depletion of intracellular guanosine triphosphate (GTP), which is required for viral RNA synthesis [Articles:11602568, 11100123]. Lastly, it has been suggested that RBV may exert its effect via immunomodulatory activity [Article:17151366].
Pharmacogenomics
IFNL3 variation and PEG-IFN-α/RBV treatment response
Marked variation in HCV clearance and response to PEG-IFN-α among different ethnic groups and among patients infected with the same HCV genotype have long pointed to host genetic factors as determinants of treatment success [Article:22641049]. In 2009, several genome-wide association studies (GWAS) independently showed that several SNPs on chromosome 19, near IFNL3, are strongly associated with response to PEG-IFN-α/RBV therapy for individuals infected with HCV genotype 1 or 4 infection. These studies demonstrated that HCV genotype 1 patients with favorable variants in the IFNL3 locus (rs12979860 CC or rs8099917 TT) were 2-to-3 times more likely to achieve SVR with PEG-IFN-α/RBV combination therapy and also had higher rates of spontaneous HCV clearance absent of treatment, as compared to patients with the unfavorable T- or C- genotypes, respectively [Articles:19684573, 19749758, 19759533, 19749757, 20060832, 19759533, 20637200, 21413051]. These findings were later validated in several candidate gene studies [Articles:20728570, 20578254, 20576307, 20176026]. The association between IFNL3 genotype and SVR has been confirmed by independent studies in various populations from Asia, Europe, and Latin America [Articles:19749757, 21911885, 22301466, 21907615, 21987611, 21346780, 22368681, 21447862, 24114761]. Thus, IFNL3 variation constitutes the strongest pre-treatment indicator of PEG-IFN-α/RBV response for HCV genotype 1 patients, even when accounting for known clinical predictive factors [Articles:19684573, 20060832]. Once PEG-IFN-α/RBV therapy is initiated, the favorable rs12979860 CC genotype is also associated with significantly higher rates of viral clearance at the 2, 4, 12, and 48-week treatment points, indicating better viral kinetics, as well as increased rates of RVR, EVR, end-of-treatment response and reduced virological relapse rate [Article:20399780].
Most studies of patients infected with HCV genotypes 2 and 3 have found relatively weak correlation between IFNL3 genotype and treatment response, but most studies failed to achieve statistical significance [Articles:21612542, 20060832, 20621700, 21112657, 21360545, 21112660], likely because SVR rates are generally high among such patients [Article:24114761]. Further, as variation in SVR rates among the different IFNL3 genotypes is generally smaller in patients infected with HCV genotypes 2 and 3, the value of IFNL3 genotyping may be of little value in predicting SVR. However, significant associations between IFNL3 genotype and RVR have been demonstrated in studies of HCV genotype 2 and 3 patients [Articles:21916985, 21254157, 21145807, 21374656]. IFNL3 genotype may be more useful in predicting RVR in patients with HCV genotypes 2 and 3, which can inform decisions to shorten treatment duration and minimize treatment-associated adverse effects [Article:24325405].
Role of IFNL3 genotype in HCV-HIV co-infected and liver transplant patients
Co-infection with HCV is relatively common (15-40%) among patients with human immunodeficiency virus (HIV), likely due to common routes of transmission [Article:17502718]. As the progression of HCV-related liver disease is accelerated in co-infected patients [Articles:11462196, 14679458], HCV has become an important cause of morbidity and mortality among HIV-infected individuals [Article:16908797]. Unfortunately, HIV-HCV co-infected patients are also less likely to respond to PEG-IFN-α/RBV therapy compared to HCV mono-infected patients [Article:17502718]. Thus, predictors of response to PEG-IFN-α/RBV treatment are useful for identifying the best candidates for current therapies. The association between IFNL3 genotype and PEG-IFN-α/RBV therapy outcome has been consistently demonstrated across studies in subgroup analyses of HIV patients co-infected with HCV genotype 1 and 4, but not in patients with HCV genotypes 3 and 4 [Articles:21612542, 21916985]. Several studies have shown an association between the favorable IFNL3 genotypes (rs12979860 CC or rs8099917 TT) and higher SVR rates [Articles:21612542, 21916985, 20389235, 20804372, 21048934], although one study that compared the two SNPs found that rs12979860 was a better predictor of response [Article:22328925]. IFNL3 genotyping may also be useful in patients co-infected with HIV and HCV genotype 1 or 4 who have previously failed PEG-IFN-α/RBV therapy in order to identify non-responders who may benefit from retreatment [Articles:1916985, 21537116].
In addition, IFNL3 genotype has been demonstrated to be an important predictor of reinfection in HCV patients who undergo liver transplantation. In several independent cohorts, the favorable rs12979860 CC and rs8099917 TT genotypes of both donor and recipient have been shown to be associated with higher SVR rates [Articles:21466653, 22411462, 22766768, 21384511]. However, most studies that have identified correlations have been performed in HCV genotype 1 patients and there is little evidence of associations among other genotypes [Article:24114761].
Distribution of IFNL3 alleles among ethnic groups
The favorable IFNL3 rs12979860 polymorphism is differentially distributed among ethnic groups, with African-Americans having the lowest frequency (allele frequency 0.39), Eastern and South-Eastern Asians the highest (nearing 0.9), and Caucasians and Hispanics having an intermediate frequency (0.63 and 0.55, respectively) [Articles:19684573, 19759533, 20399780, 22438096]. The SNPs rs12979860 and rs8099917 exhibit strong linkage disequilibrium except in patients of African ancestry, with partial linkage disequilibrium in Caucasians and near-complete linkage disequilibrium in Asians [Articles:19684573, 20060832]. The varying frequencies of favorable alleles help to explain differences in SVR rates among different ethnic populations [Articles:19684573, 19826275]. For example, it is estimated that approximately half of the difference in SVR rates among European-Americans and African-Americans is due to the rs12979860 C allele [Article:19684573]. Still, in African-American patients with the favorable rs1297860 CC genotype, viral kinetics have been shown to be slower and SVR rates lower than in European-Americans of the same genotype. This may suggest that ethnicity remains an independent predictor of outcome and pointing to the existence of currently unknown gene variants that influence treatment response, particularly in African-Americans [Article:20399780].
Proposed mechanism of IFNL3 variations on treatment response
The biological implications of IFNL3 variations, the actual mechanisms by which IFNL3 genotype affects response to PEG-IFN-α and RBV, remain unclear. While the effects of IFNL3 polymorphisms on IFN-λ production and intrahepatic ISG expression are still controversial, it has been suggested that ISG expression patterns may in part explain the difference in SVR rates. Interestingly, patients with the unfavorable IFNL3 genotype (rs1297860 CT/TT) were found to have higher intrahepatic ISG expression at baseline and this is associated with failure to respond to PEG-IFN-α/RBV treatment [Articles:18467494, 20434452]. One explanation for this is that patients with the unfavorable genotype exhibit continual, ineffectual intrahepatic expression of ISGs. At the same time, IFN-signaling inhibitors such as SOC3 and PIAS1 are also upregulated; this pre-activated ISG response may be insufficient to clear the virus while causing reduced sensitivity to PEG-IFN-α therapy through negative regulation of JAK-STAT signaling by IFN-signaling inhibitors [Articles:18467494, 21898478]. Thus, even when PEG-IFN-α is administered, the cell may fail to induce strong enough ISG expression and be unable to clear the virus [Article:21898478].
Conversely, patients with the favorable IFNL3 genotype may induce a weaker response to HCV RNA and thus less IFN expression occurs, resulting in lower ISG levels at baseline and higher viral loads. When IFN-α is administered, there may then be less inhibition of IFN-signaling by negative regulatory molecules, causing the cell to be more sensitive to IFNs, and a stronger ISG induction is possible. Ultimately, this unrestrained IFN signal transduction and strong ISG stimulation may result in more effective clearance of the virus. Indeed, patients with the favorable IFNL3 genotype are observed to eliminate HCV more efficiently at each time point of treatment, indicating better viral kinetics [Article:20931559]. Yet, despite known association with treatment response and a proposed mechanism for its effect, it remains unknown whether the favorable IFNL3 SNPs actually exert biological effects or are simply in linkage disequilibrium with other functional polymorphisms.
IFN4 (ss469415590) and PEG-IFN-α/RBV treatment response
Most recently, studies of the genetic region upstream of IFNL3 have uncovered a new transiently induced region that encompasses a dinucleotide variant, rs368234815 TT/ΔG (originally designated ss469415590), which is in high linkage disequilibrium with rs12979860 and has been suggested as a possible causal variant [Articles:24786669, 23291588, 23712427]. ss469415590[ΔG] is a frameshift variant that creates a novel gene, IFNL4, encoding the IFN-lambda4 protein (IFN-λ4). IFN-λ4 is similar but not identical to IFN-λ3 [Article:23291588]. Like the existing three members of the IFN-λ family, IFN-λ4 exhibits antiviral activity in vitro and binds the IL10RB/IFNLR1 receptor complex to activate the JAK-STAT pathway. Patients with the ss469415590[ΔG] allele express the full protein, while the TT allele causes a frame shift that leads to an abrogation of the IFN-λ4 protein [Article:23291588]. Interestingly, the TT allele is strongly associated with spontaneous or treatment-induced SVR, indicating that disruption of IFNL4 is favorable [Articles:23291588, 23712427]. Although the mechanism remains unclear, the apparent associations between IFN-λ4 production and a reduced likelihood of spontaneous or PEG-IFN-α/RBV therapy-induced clearance of HCV suggests this novel IFN may be responsible for the observed clinical phenotypes [Article:23291588]. In contrast to most IFNs, IFN-λ4 protein is poorly secreted and it has been hypothesized that it may impair HCV clearance by blocking other IFN-λs from binding the IL10RB/IFNLR1 receptor. Of note, ss469415590, but not rs12979860, has been shown in peripheral blood mononuclear cells to be involved in the stimulation of IFN-α and IFN-γ-inducible protein 10 (IP-10) mRNA, high plasma levels of which are associated with treatment failure [Article:23712427].
As ss469415590 is strongly correlated with rs12979860 in IFNL3, the novel variant provides no additional information for predicting treatment response in Caucasian patients. Conversely, IFNL4 is only moderately correlated with rs8099917 in Caucasian patients, suggesting it may be a better predictor of treatment outcome [Articles:24205831, 24308755]. Likewise, IFNL4 genotype may be a better predictor of treatment response in patients of African ancestry as ss469415590 shows only weak correlation with rs8099917 and moderate correlation with rs12979860 in these populations [Article:23291588]. Taken together, the identification and characterization of the ss469415590[ΔG] functional variant represents a new step towards elucidating genetic mechanisms of HCV clearance and the role of IFN polymorphisms in predicting HCV treatment response.
IFNL3 in the age of DAAs
With the incorporation of DAAs into the HCV treatment arsenal, the question emerges of whether IFNL3 genotype will remain relevant in predicting treatment outcomes to triple acting therapies that include these new agents. Initial data suggest that IFNL3 genotype may remain useful, but less so than for traditional PEG-IFN-α/RBV dual therapy [Articles:21628662, 20648473]. To date, the most well-studied triple therapies are those which combine PEG-IFN-α and RBV with the 'first generation' protease inhibitors BOC and TVR. In treatment naïve patients, SVR rates for patients treated with triple therapies that included BOC or TVR, respectively, were significantly higher for those with the favorable rs12979860 CC genotype (80% and 90%) as compared with CT (71%, and 71%) or TT (59% and 73%) [Articles:21696307, 21449783, 22626609]. Further, patients exhibited better viral kinetics of response and a lower risk of selection of resistance-associated HCV variants [Articles:22626609, 23321318]. In patients previously treated with PEG-IFN-α/RBV therapy, IFNL3 genotype was not significantly associated with SVR in those who had relapsed or in partial responders, but previous null responders with the favorable rs12979860 CC genotype exhibited higher SVR rates compared to those with unfavorable genotypes [Article:20375406].
With regard to the latest developed 'second generation' protease inhibitors, the effect of IFNL3 genotype on treatment response has been shown to be weaker but is expected to remain useful in predicting response, especially in treatment naïve patients. For example, IFNL3 genotype has been shown to be significantly associated with SVR at week 12 in both treatment-naïve and treatment experienced HCV genotype 1 patients undergoing triple therapy with the NS3 protease inhibitor simeprevir [Article:24907225]. Similarly, rs12979860 was identified as an independent pre-treatment predictor of SVR in treatment-naïve patients with HCV genotypes 1, 4, 5 and 6 treated with the NS5B polymerase inhibitor sofosbuvir in combination with PEG-IFN-α/RBV [Article:23944316]. It has also been suggested that stratification of patients according to IFNL3 genotype in on-going clinical trials of new treatments will be useful in individualizing future triple therapies and optimizing outcomes [Article:25114601]
Lastly, IFNL3 appears to marginally affect treatment response in newly developed IFN-free therapies [Article:22248659]. While IFNL3 genotype may not be relevant in predicting SVR, it has been demonstrated that IFNL3 variations are associated with viral kinetics during IFN-free treatments [Articles:22248659, 23944300]. However, studies with larger cohorts will be needed to confirm these associations.
Other host genes and PEG-IFN-α/RBV response
Apart from the well-established associations described above, numerous studies have demonstrated that polymorphisms in other genes may be associated with treatment response and adverse reactions to PEG-IFN-α/RBV therapy. One of the most significant drug-related toxicities is RBV-induced hemolytic anemia, which causes approximately 15% of patients to reduce RBV dose or discontinue treatment [Article:20652658]. In 2010, a GWAS identified a polymorphism of the ITPA gene (rs6051702 A), encoding inosine triphosphate pyrophosphohydrolase, shown to protect against ribavirin-induced hemolytic anemia in HCV genotype 1 patients treated with PEG-IFN-α/RBV [Article:20173735]. Subsequent analysis identified SNPs on two functional variants carrying the rs6051702 C allele that also protect against RBV-induced anemia in an European-American HCV genotype 1 population [Article:10733558]. Further, these results have been replicated in a Japanese population and extended to HCV genotype 2, 3 and 6, as well as patients treated with PEG-IFN-α/RBV plus TVR triple therapy [Articles:20637204, 20547162, 22584257, 23730840, 21246582, 23707372]. While these variants have established associations with ITPA deficiency, the mechanism by which this condition protects against RBV-induced anemia remains unclear.
A genetic variant of the LDLR gene, which encodes the low-density lipoprotein (LDL) cholesterol receptor, has been shown to predict SVR in chronic HCV patients treated with PEG-IFN-α/RBV therapy [Article:12209363]. LDLR and IFNL3 variants appear to have additive effects on predicting SVR [Article:21572301]. Thus, consideration of both LDLR and IFNL3 is more accurate in predicting SVR than use of either genotype singly. Lastly, it has been shown that 25-OH vitamin D deficiency is associated with nonresponse to PEG-IFN-α/RBV combination therapy, suggesting that variants of the vitamin D receptor gene (VDR) may play a role in treatment outcomes [Articles:21145801, 15315818]. It was recently demonstrated that a common nonsynonymous SNP in the VDR gene (rs2228570) predicts treatment response to PEG-IFN-α/RBV therapy for HCV genotype 1 and 4 patients, with minor T allele carriers having an increased likelihood of achieving SVR [Article:24073221].
Conclusion
Treatment of HCV genotype 1 infection has made remarkable strides over the last decade, allowing SVR rates to climb to new highs for a chronic infection that has been traditionally difficult to cure. In treatment naïve patients, an approximate 30% increase in SVR has been possible with the addition of DAAs to PEG-IFN-α/RBV combination therapies. Pharmacogenetics offers a useful tool to optimize individual treatment regimes by allowing clinicians to make more informed decisions on treatment options, modify dose and duration of treatment regimes and avoid certain adverse reactions associated with therapies. In particular, IFNL3 genotype has been established as a robust predictor of treatment outcomes to PEG-IFN-α/RBV-based therapies. In the era of DAAs, IFNL3 variation will likely remain useful, although the effects will be attenuated. However, newer therapies are extremely expensive and thus PEG-IFN-α/RBV-based regimes will likely remain the mainstay of HCV treatment in developing nations. As developed nations like the U.S. adopt newer, more effective treatments, studies will be needed to confirm initial associations found between IFNL3 genotype and treatment response. Although likely theories have been suggested, it is still not certain how IFNL3 genotype affects treatment outcomes and further research will be needed to elucidate the exact mechanism. Improved mechanistic understanding of PEG-IFN-α and genetic determinants of SVR will help to identify novel biomarkers for spontaneous clearance, determine new targets to personalize treatments for HCV infection and ultimately allow for further improvement of treatment response rates. As SVR rates climb to new highs of greater than 95% with the advent of IFN-free regimes, which have now replaced IFN-based therapies in developed nations, the future of genotype 1 HCV treatment is bright indeed.
Edit history (1)
- 2024-06-13 Update fixed typos